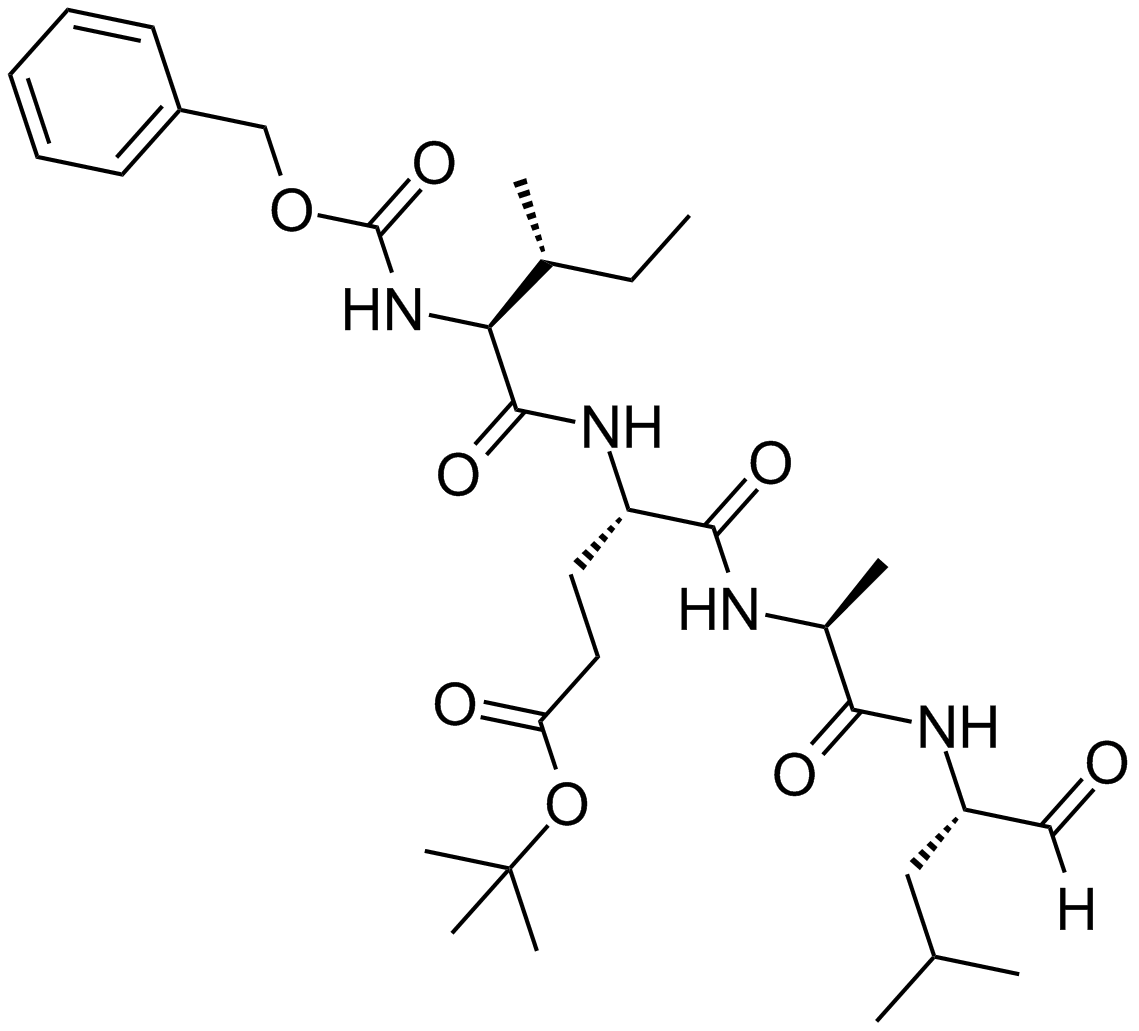
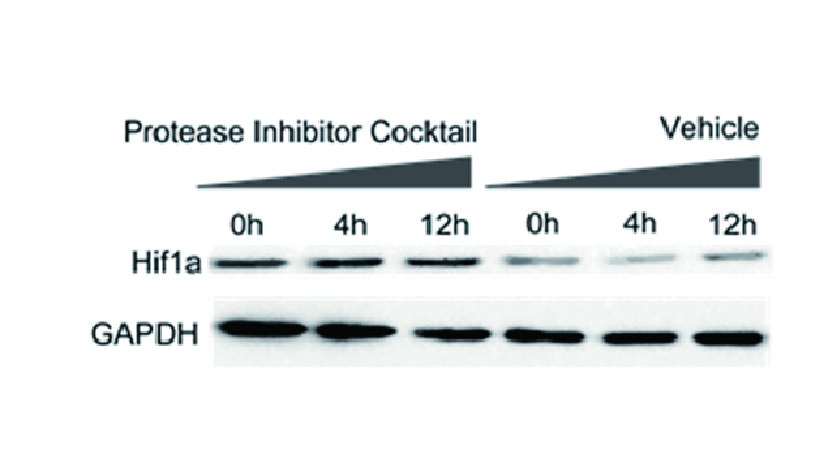
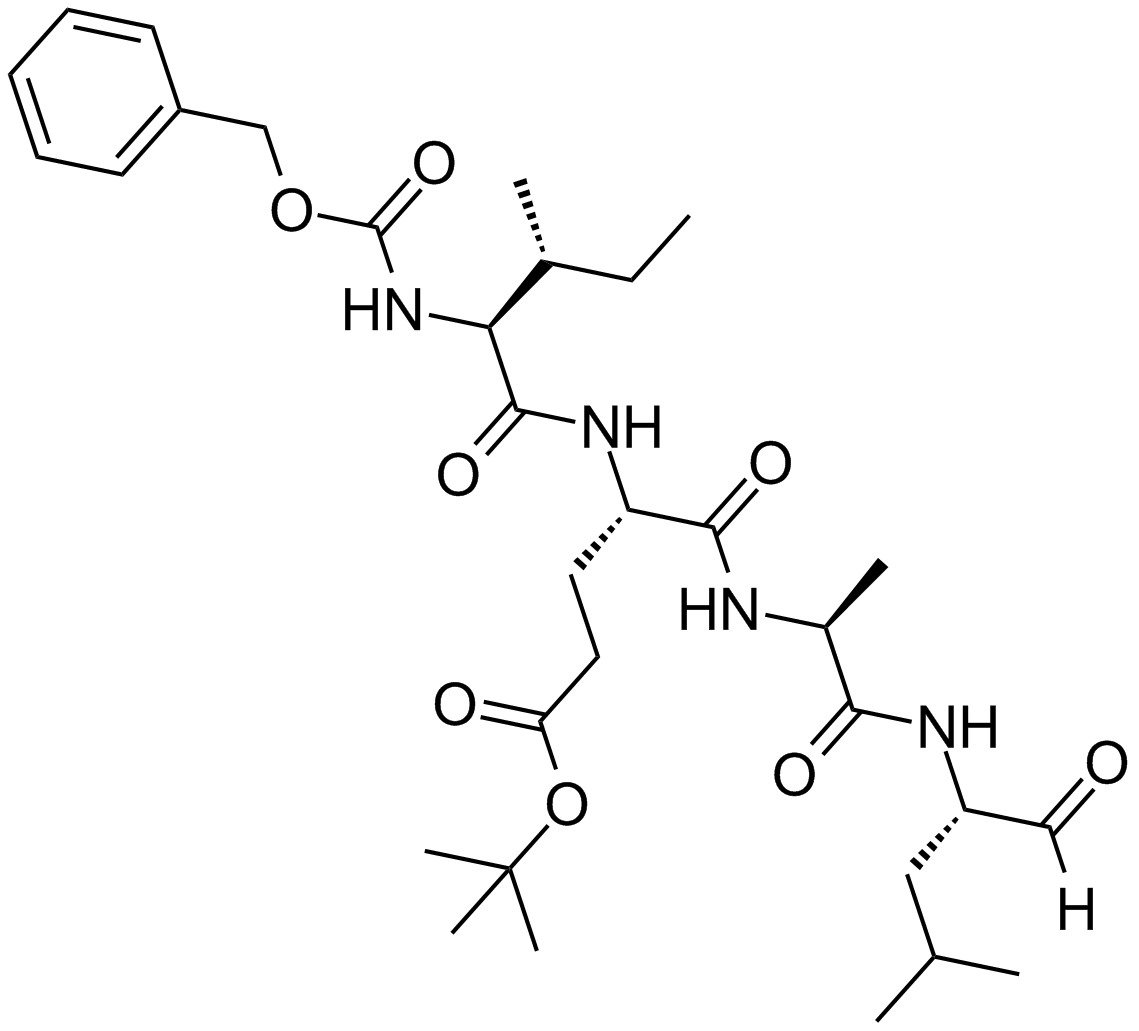
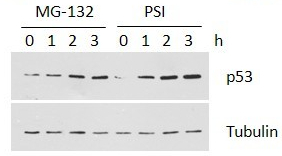
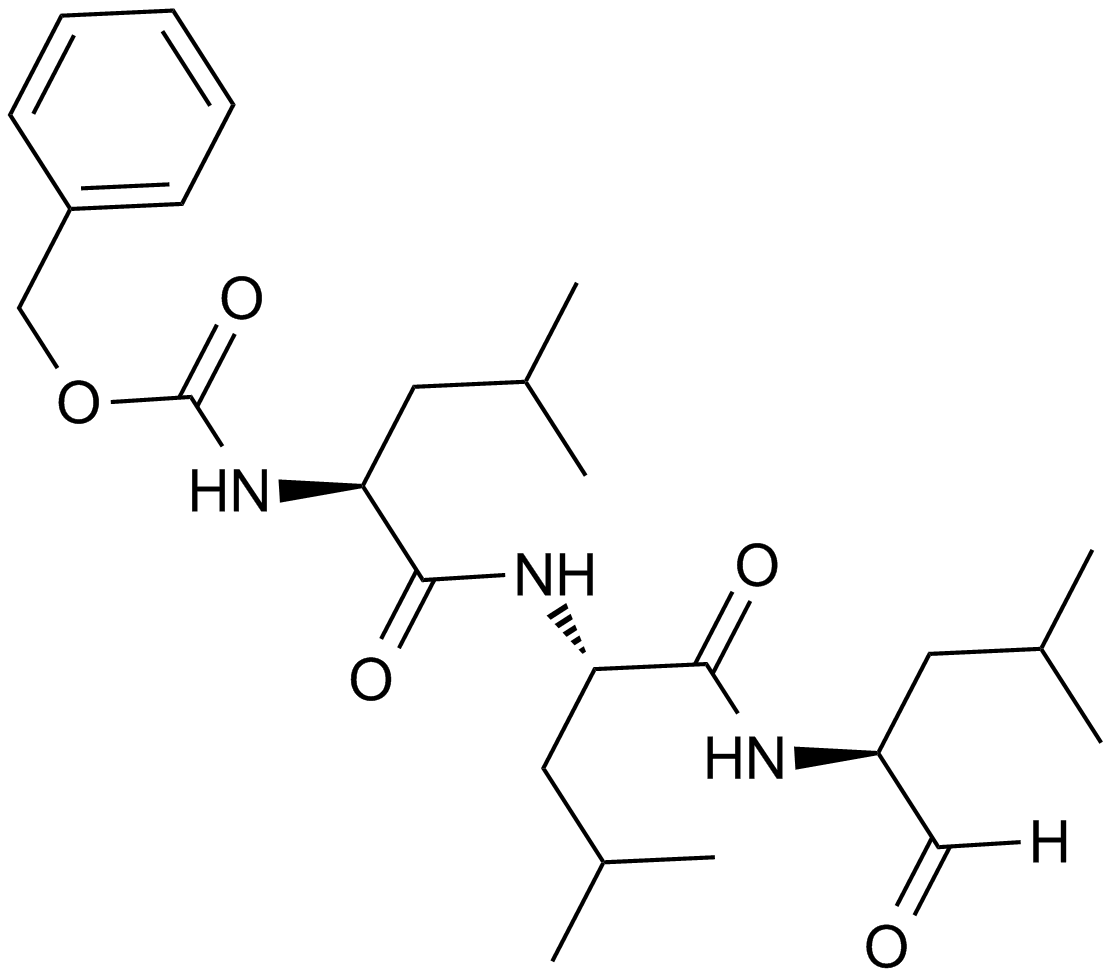
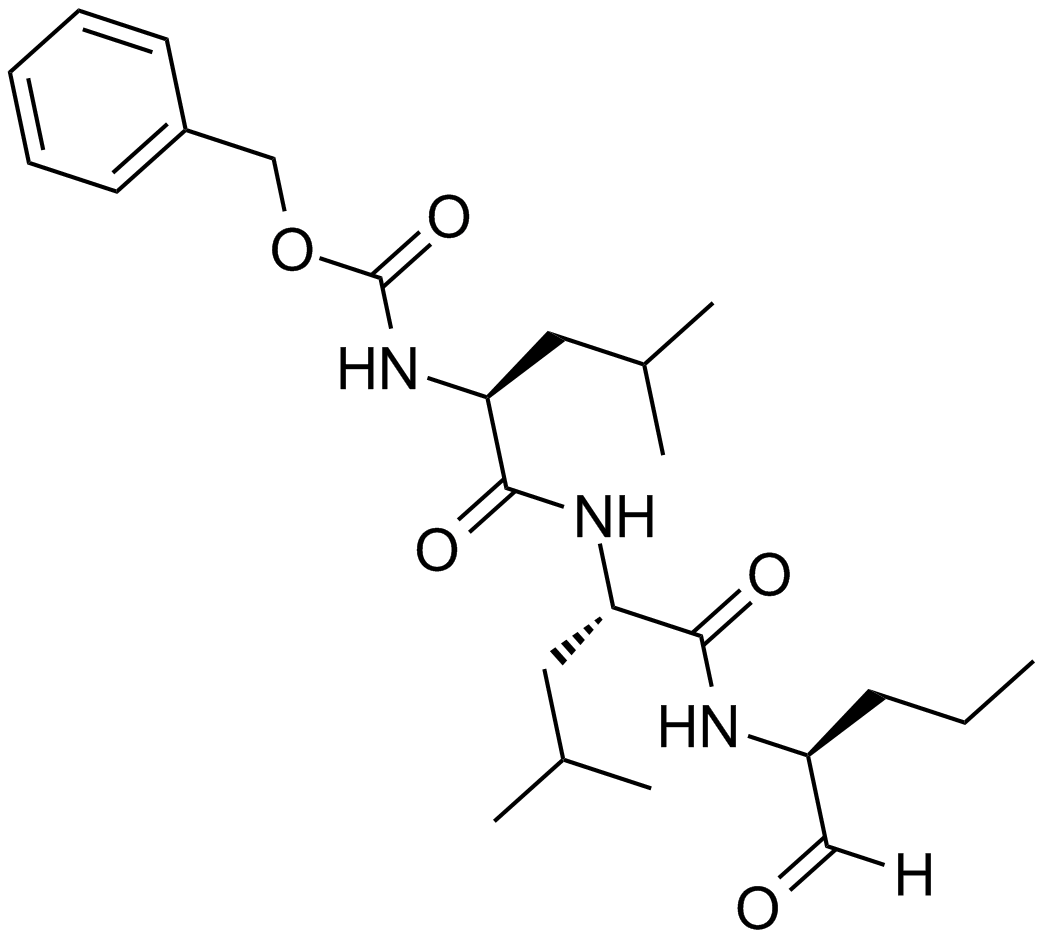
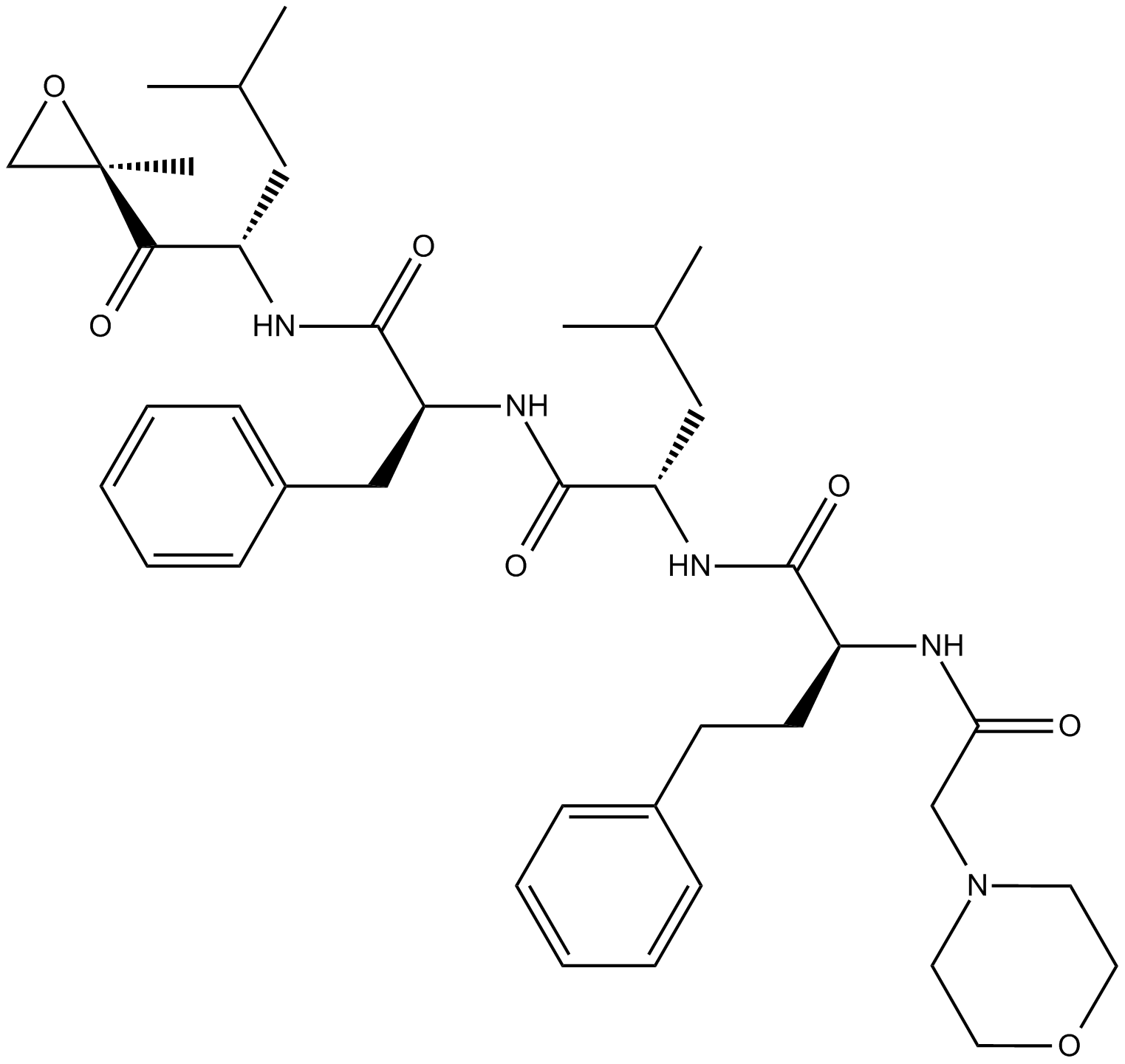
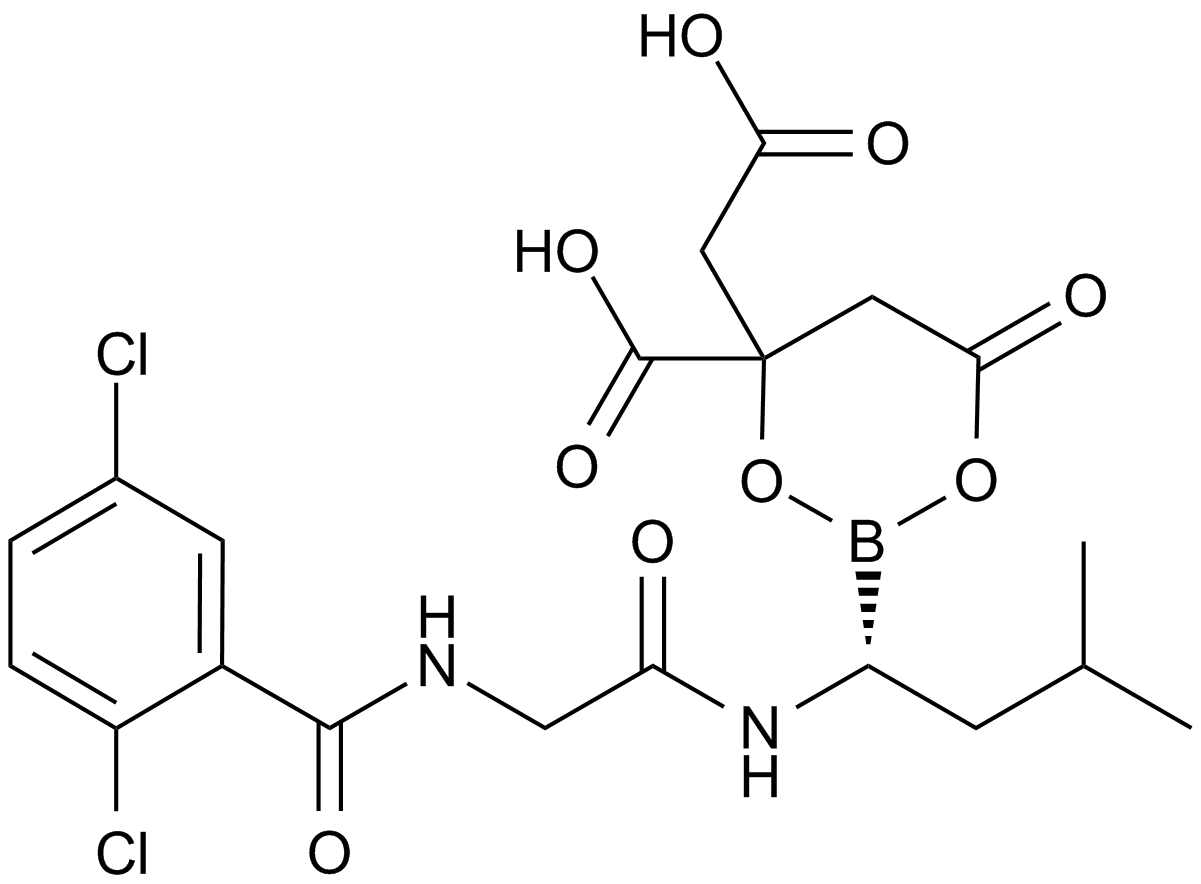
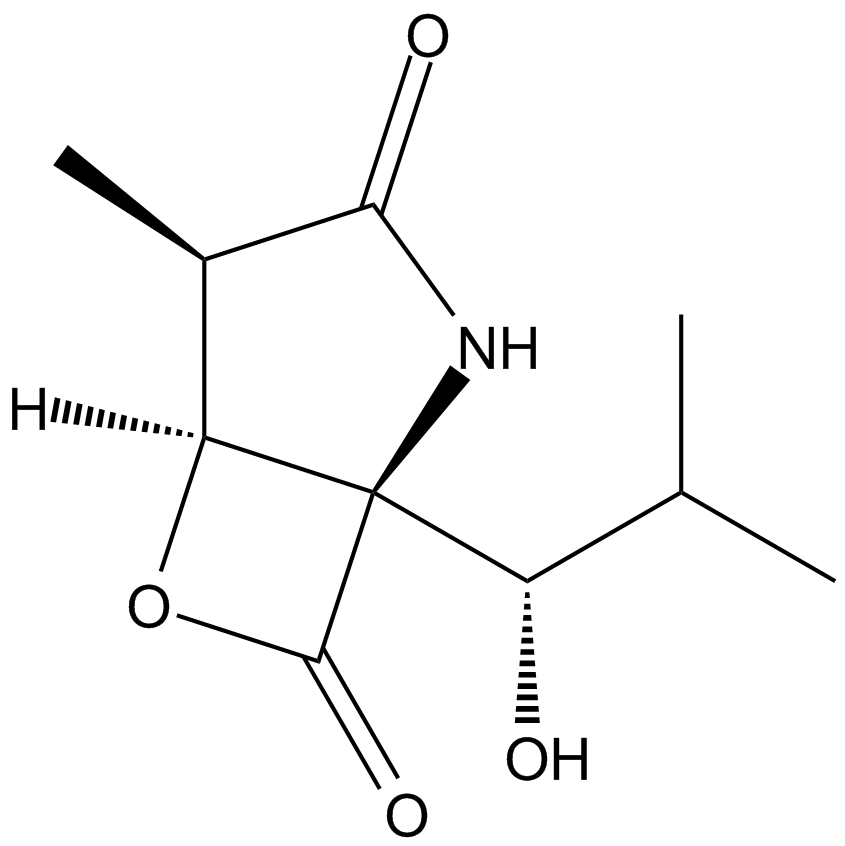
ZIE(OtBu)AL-CHO (PSI)1 have been shown to inhibit the proteasome activities in a variety of cell types.
Peptide aldehyde, PSI (Z-Ile-Glu(OtBu)-Ala-Leu-al), inhibits the proteasome 10-fold better than calpain but is less potent than MG1322. Since MG132, PSI, MG115 (Z-Leu-Leu-nVal-al) and ALLN can all inhibit calpains and various lysosomal cathepsins in addition to the proteasome, when using these inhibitors in cell culture it is important to perform control experiments to con¢rm that the observed e¡ects are due to the inhibition of the proteasome. First, one can use agents, which block intracellular cysteine proteases, but do not inhibit proteasomes3. Such inhibitors are Z-Leu-Leu- al, and E-64 for calpains4, and weak bases such as chloroquine and E-64 for lysosomal proteolysis . In yeast, where digestive vacuoles contain mainly serine, not cysteine, proteases, phenylmethylsulfonyl £uoride can be used to inhibit these enzymes without affecting proteasomes5.
Despite the availability of these inhibitors, MG132, due to its low cost and the rapid reversibility of its action, still remains, in our opinion, the first choice to study proteasome involvement in a process in cell cultures or tissues, if appropriate controls are used. As the most potent and selective of commercially available aldehydes, MG132 is preferable to ALLN, MG115 (Z-Leu-Leu-nVal-al), or even PSI. On the other hand, the least selective inhibitor, ALLN, because of its ability to inhibit most major pro teases in mammalian cells, is probably the best tool for prevention of unwanted proteolysis, for example during isolation of proteins from mammalian cells.
Reference:
1. Takada K (1995) Mol. Biol. Rep. 21: 21–26
2. A. F. Kisselev, A. L. Goldberg. Proteasome inhibitors: from research tools to drug candidates. Chemistry & Biology 8 (2001) 739-758.
3. W. Matthews, J. Driscoll, K. Tanaka, A. Ichihara, A.L. Goldberg, Involvement of the proteasome in various degradative processes in mammalian cells, Proc. Natl. Acad. Sci. USA 86 (1989) 2597-2601.
4. S. Tsubuki, Y. Saito, M. Tomioka, H. Ito, S. Kawashima, Differential inhibition of calpain and proteasome activities by peptidyl aldehydes of di-leucine and tri-leucine, J. Biochem. 119 (1996) 572-576.
5. D.H. Lee, A.L. Goldberg, Selective inhibitors of the proteasome-dependent and vacuolar pathways of protein degradation in Saccharomyces cerevisiae, J. Biol. Chem. 271 (1996) 27280-27284.